Let’s build on the convergence between vertebrates and cephalopods. This time, let me quote some excerpts from the following article: Squid vascular endothelial growth factor receptor: a shared molecular signature in the convergent evolution of closed circulatory systems by Masa-aki Yoshida, Shuichi Shigeno, Kazuhiko Tsuneki, and Hidetaka Furuyaa (in EVOLUTION & DEVELOPMENT 12:1, 25–33 (2010)).
Let’s build on the convergence between vertebrates and cephalopods. This time, let me quote some excerpts from the following article: Squid vascular endothelial growth factor receptor: a shared molecular signature in the convergent evolution of closed circulatory systems by Masa-aki Yoshida, Shuichi Shigeno, Kazuhiko Tsuneki, and Hidetaka Furuyaa (in EVOLUTION & DEVELOPMENT 12:1, 25–33 (2010)).
First, the researchers lay the groundwork for the convergence of these two systems:
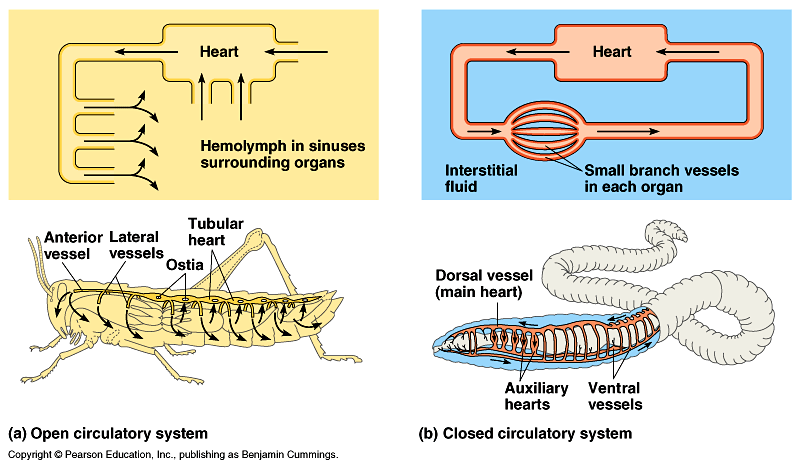
Metazoan animals have evolved an incredible diversity of hearts and heart-like structures. The most elaborate case in invertebrates is observed in coleoid cephalopods: they exhibit an elaborate closed circulatory system (Schipp 1987; Budelmann et al. 1997). Their heart possesses a kind of advanced output structure similar to that of the human heart, which differs largely from molluscan typical nonendothelium primitive chambered hearts (see Kling and Schipp 1987; Schipp 1987). Neither morphological nor molecular data give strong support to a close phylogenetic relationship between vertebrates and cephalopods, suggesting that the closed circulatory systems and complicated hearts were formed independently in each lineage, and have converged during their evolution.
and
Their cardiovascular system is considerably similar to vertebrates in several respects such as high oxygen binding capacity, high concentrations of proteins, and short circulation time (Schipp 1987). Each vessel in the cephalopod is constructed similarly to vertebrate vessels, with an endothelial lining on a basement membrane (Budelmann et al. 1997), although the cephalopod blood vessel lining does not have the cellular junction typical among vertebrate species. Most invertebrates have no endothelium in their vascular walls so the cephalopods are unusual in that they are invertebrates with vertebrate type blood vessels. As the other molluscs have open vascular system, the peculiar blood vessel configuration in the cephalopods is in all probability secondarily developed similarly to the vertebrate among chordates (Ruppert and Carle 1983).
Then, they find something really cool.
Continue reading →